

In the struggle for survival, predators need to capture and consume prey, and prey have to avoid being eaten. Over evolutionary time this has evolved into a kind of arms race between the two, in which predators have evolved specialised weapons of attack and prey have evolved specialised defences. One particular example of this is the arms race between bats and moths.
The interaction between bats and their insect prey, in particular moths, is one of the most cited examples of such an evolutionary arms race. It comes with a twist – the weaponry used by each is largely based on sound and hearing.
Evolving attacks and defensive systems
The evolutionary arms race between bats and moths was initiated by bats evolving echolocation, which allowed them to detect moths in complete darkness. Echolocation is a form of biological sonar. Bats emit high frequency calls and frequency is the number of waves of sound generated per second. They locate prey by listening for the echoes generated when their calls are reflected off the prey.
The frequencies used by bats ranges from 12–210 kHz. Since human hearing disappears at about 20 kHz, this means that most bats echolocate at frequencies much higher than humans can hear.
Moths developed an early warning system to defend against the new way bats attack. Moths have evolved ears which allow them to hear bat echolocation, execute evasive flight manoeuvres and avoid being eaten. Some bats in turn improved their offensive weaponry by evolving stealth echolocation – that is, echolocation at frequencies and intensities that are inaudible to moths. Some moths, already equipped with ears, have in turn evolved ultrasonic clicks of their own that are used defensively during a bat attack.
Evidence that moth ears have evolved in the context of bat predation is provided by the close match between the frequencies at which moths hear best and the frequencies of the calls of bats that prey on them.
In North America, most insect-eating bats echolocate between 20-50 kHz. This is also the range at which moths hear well. In Africa and Australia – where moths are preyed on by bats that use echolocation frequencies higher than 50 kHz – the upper hearing range of moths can extend to 100 kHz and beyond.
Moths also use more active forms of defences in the form of ultrasonic clicks. Moths produce these clicks using tymbal organs located at the front of the thorax just behind the head. Bats break off their attack on moths when the moths emit their high-frequency clicks. Three explanations for how moth clicks work have been proposed: clicks startle the bat forcing it to abort its attack, clicks alert the bat to the moths’ unpalatability and moth clicks jam bat echolocation by interfering with the bat’s ability to process returning echoes.
These adaptions have taken place over a million years. All players in predator/prey interactions will evolve adaptations and counter adaptations.
The bat variations
Bats may counter the auditory defences of moths by reducing their acoustic conspicuousness in several ways, including using frequencies which moths can’t hear and/or reducing the intensity and duration of their calls. There is evidence that some species of bats may have evolved echolocation frequencies that are inaudible to moths.
For example, the Spotted bat in North America uses particularly low frequency calls (12 kHz), and the Short-eared trident bat in Africa uses an unusually high call frequency (208 kHz). Both eat moths almost exclusively.
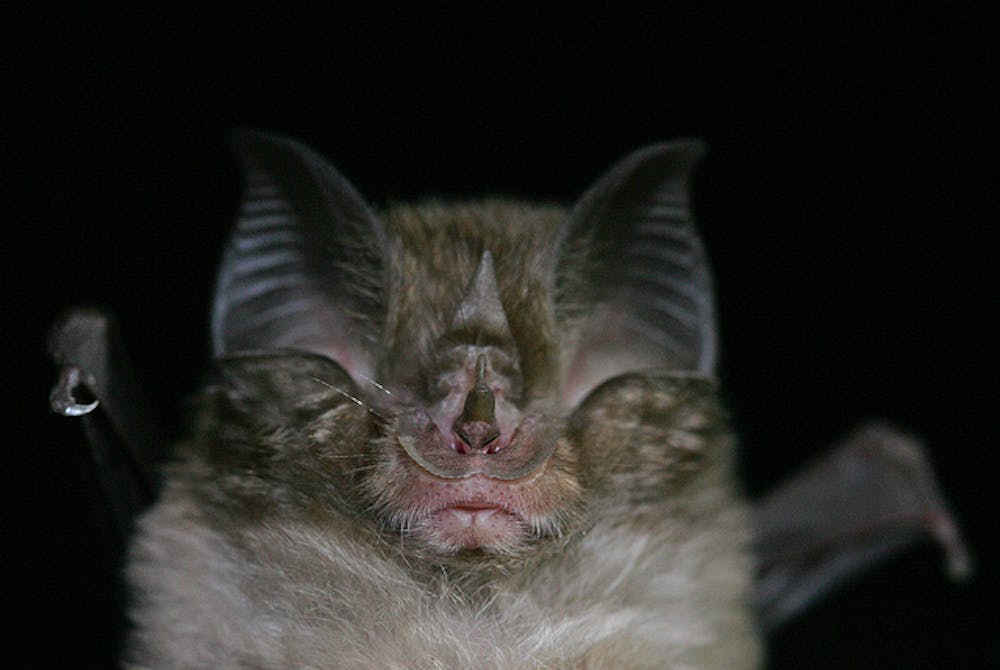
Horseshoe bats and Old World leaf-nosed bats use very high echolocation frequencies and feed predominantly on moths, suggesting that they are able to successfully prey on them because their high call frequencies are inaudible to moths.
However, such associations on their own cannot be used as definitive support for the notion that high call frequencies in bats have evolved because they are less audible to moths. We have to show that these calls are in fact inaudible to moths. The results are mixed.
Along the south coast of South Africa, the calls of the Cape horseshoe bat echolocating at 80-86 kHz are audible to the most common moths that co-occur with this bat. Conversely, however, most of the calls of the Dusky leaf-nosed bat (calling at 50–160 kHz) were inaudible to co-occurring moths. In some cases, moths are ahead in the arms race and in some cases bats are.
Audibility of a sound is dependent on frequency and intensity, and so bats could also lower the intensity of their calls to make them less audible to moths. The Barbastelle bat in Europe appears to have done just that. The echolocation of this species is of much lower (10 to 100 times lower) intensity than the calls of other bats which hunt in the same way.
As a consequence of its lower intensity calls, moths are unable to detect this bat until it is too late to execute effective evasive manoeuvres. The effectiveness of the low intensity calls of the Barbastelle bat in overcoming the hearing defence of moths is supported by it feeding almost exclusively on eared moths.
As we continue our research into this endless evolutionary dance between bats and moths, we are likely to uncover even more amazing adaptations and counter-adaptations.