

Wisdom teeth, the palmaris longis tendon, ear wiggling: these qualities were desirable millions of years ago, but due to changes in our diet and environment, are slowly disappearing. However, such features aren’t just useless remnants - they provide valuable insights into our evolutionary past.
One of Charles Darwin’s most enduring intellectual legacies was the idea of “common descent” – that the history of life is like a tree with different evolutionary lines sprouting from a common trunk.
Put another way, he recognised that all of life was traceable to a single common ancestor, with evolutionary history unfolding through the inheritance of common features, modified through time as organisms evolved in response to changes in their environment.
As we pass back in time, we humans, as a species, must have shared thousands of common ancestors with other organisms over the past three billion years or more of the history of life none of which we will ever firmly identify.

It begins for us with the Neandertals – our closest extinct cousin – onto chimpanzees – our closet living relatives – and ultimately back through the tree of life of primates, early mammals and even earlier animals back to the very earliest life when we shared a common ancestor will all living things.
For 150 years or so, biologists have studied the human body for the vestiges of our evolution in the form of apparently redundant features dubbed “vestigial” organs.
This evolutionary “excess baggage” has often been thought to reflect common descent and sometimes even stages of evolution. While our views of vestigial features have changed in that time, they are still informative about both our past and the genetic makeup of our species.
Evolutionary hangovers
Since Darwin’s time, students of natural history have been fascinated by the presence of apparently non-functioning (vestigial) human body structures as evidence for our evolution.
Dubbing them vestigial assumes of course an absence of function, or perhaps disuse. Rather, this turns out to have been wrong in almost all cases, reflecting our ignorance as scientists rather than evolution’s inertia.
When it comes to the fossil record – where function is so much harder to determine – the debate is normally over whether ancient features truly reflect the lifestyle of an organism or whether they might simply be evolutionary excess baggage.

This is an important issue in human evolutionary science. For example, the skeletons of many australopithecines (such as “Lucy”) possess a mix of features associated with both ape-like tree climbing and human-like terrestrial (bipedal) locomotion.
An important and difficult question to answer in Lucy’s case is: do the tree climbing features in her skeleton reflect evolutionary hangovers – inherited primitive features that are not functionally important – or are they indicators of a life partially lived in the trees? In all honesty, we’re really not sure.
In his book The Descent of Man, first published in 1871, Darwin listed a large number of human “parts in a rudimentary condition”, being body structures that are either “absolutely useless” or “they are of such slight service” owing to the fact that the conditions under which they evolved no longer exist.
Some interesting examples are:

Smell or olfaction: while humans, being primates, rely heavily on vision, our sense of smell is actually quite well developed with more than 300 genes associated with this special sense. The human nose is even able to detect concentrations as low as a few parts per trillion for some odorous compounds.
Our sense of smell is clearly and intimately linked to our sense of taste – with perhaps 80-90% of taste attributable to smell – plays a role in our social interactions, including our choice of mates, and helps keep us safe by alerting us to hazards in the environment.
Body hair: our near nakedness remains one of the greatest mysteries of human evolution. One suggestion is that it may have been associated with endurance running in our evolution 2-3 million years ago owing to the increased convection rates associated with hair loss. This leaves open the question of why have body hair at all?
The meagre hair we have probably performs one or more functions. Suggestions mostly centre on a role in sexual signalling and mate choice owing to the large increases in hair growth associated with sexual maturity from puberty onwards.
Moreover, apocrine sweat glands, located in regions of the body that grow hairs with sexual maturity (such as armpits and groin), produce fluids from puberty onwards that contain pheromones, chemicals designed to elicit a change in the physiology or behaviour of another person.
Wisdom teeth: primates almost universally possess three molars on each side of their jaw, in both the top and bottom dental rows. Living humans are unusual in that one or more third molar - what we call wisdom teeth - quite commonly doesn’t erupt or fails to develop.

There is a wide range of causes for this phenomenon, including small jaws and dental crowding, but congenital factors seem to be most prevalent. While absence does occur in other primates, it seems to be much more common in humans, with population estimates frequently around 20% (versus less than 5% in other primates). Missing third molars have also been documented in low incidence in other hominins such as Homo erectus.
Reduction in third molar number may indeed be an example of a recent human evolutionary trend and seems to distinguish us from other primate species. Whether it qualifies as vestigial is a matter of perspective, but without doubt the modern agricultural/industrial diet places few demands on our teeth and combined with modern dentistry we can get away with fewer teeth.
Vermiform appendix: the human appendix was for a long time thought to be quintessential vestigial organ. It is now known to play a role in the immune system, contains lymphatic vessels that help regulate pathogens and assists with the management of digestive system movement and removal of body waste.
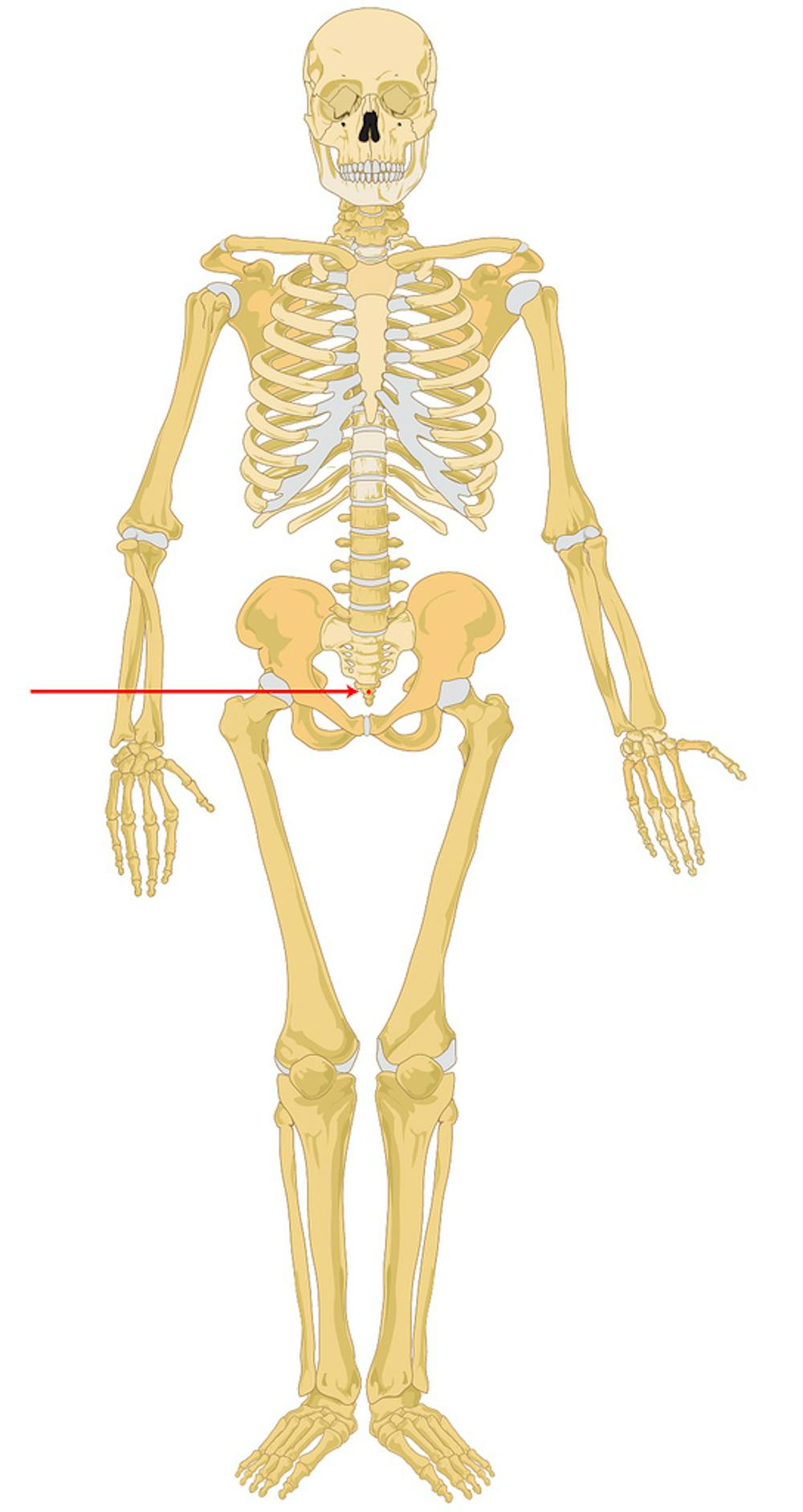
Coccyx: while our tailbones don’t bear any direct body weight when we’re upright – unlike the remaining spine and pelvis – they do play a vital role in the human body. Importantly, they provide attachment for muscles that support the pelvic organs – such as the pelvic sling muscle the pubococcygeus – as well as the major locomotor muscle, the largest of the “glutes”: the gluteus maximus.
Prostate gland: the function of this male organ, one of the accessory sexual glands, is now well understood – unlike in Darwin’s day. It makes a fundamental contribution to semen, both in the form of supporting reproduction and aiding sperm survival, and also contains antimicrobial components that may help project against urethral infections.
Vomeronasal organ: many mammals possess a pair of organs on either side of the nasal septum on the floor of the nasal cavity used for the detection of pheromones. Until recently, they were thought to absent from humans, but have now been shown to be present in most people.
While understanding of their precise function and effects remains controversial, it seems likely that, as with other mammals, their role is at least in part to detect pheromones from other people and to help regulate aspects of reproductive physiology and behaviour.
In other words, they seem to detect the pheromones produced by the apocrine sweat glands of other people that influence our decisions about mate choice and the sexual partners we select.
A true vestigial structure?
When humans walk, one foot bears the body weight while the opposite one pushes off to propel the body. The pushing off foot is normally rigid – kept so by ligaments and muscular tendons – in order to act as an efficient lever.
In other primates this foot is much more mobile and actively alters position to accommodate uneven surfaces. One way this occurs is through the “mid-tarsal break” or a bending of the mid-foot.

The mid-tarsal break is thought to have been lost in humans as an adaptation to our bipedal locomotion. Indeed, it seems to be lacking in most species of Australopithecus including Lucy’s kind, but may be present in Australopithecus sediba.
A recent study published in the American Journal of Physical Anthropology shows in a sample of 398 adult subjects studied, almost 10% of them actually possess the primate mid-tarsal break.
This runs counter to orthodoxy and is not easily explained. While the feature is associated with people with flatter arches and a tendency to roll the foot inwards when walking, the bending is counteracted with shoes, so has little apparent real impact on movement.
The mid-tarsal break, more than any other feature studied over the past 150 years of evolutionary biology, probably justifies the appellation “vestigial”. It seems to be about as compelling an example of evolutionary excess baggage as one can imagine.
Yet, further research may show even in this case that evolution has co-opted the structure for two-footed working in a way that was unexpected. It may even prove to be advantageous rather than simple inertia.
At a minimum, it powerfully shows our common descent with other apes, with whom we share the vast majority of our DNA and so many of our physical and behavioural features. We are, after all, just a “jumped-up” ape!