


Les cellules, souvent considérées comme les unités élémentaires du vivant, sont des entités déjà extrêmement complexes. Imaginer l’origine de la vie, c’est imaginer l’émergence naturelle de cellules semblables, bien plus simples que celles que nous connaissons aujourd’hui, mais quand même relativement complexes. Cela revient à dessiner un chemin tortueux qui part d’un monde purement chimique et minéral, et intègre petit à petit l’apparition d’une multitude de composés et de dynamiques. Malgré l’abondance d’expériences et d’hypothèses, aucun scénario complet d’abiogenèse (autre nom de l’origine de la vie) ne parvient aujourd’hui à convaincre la communauté scientifique internationale. Assembler tous ces travaux en des scénarios cohérents et comparer efficacement leur plausibilité reste un défi majeur.
Notre récente étude publiée dans la revue iScience pointe l’intérêt du concept de seuil pour décomposer les scénarios d’origine de la vie en une succession d’étapes formalisées, pour faciliter leur évaluation et leur ajustement. Un seuil est la limite entre deux états qualitativement distincts d’un système physico-chimique. Il est franchi lorsqu’un tel système subit un changement qualitatif majeur, à la suite d’une faible variation de paramètres de contrôle relevant du système lui-même ou de son environnement.
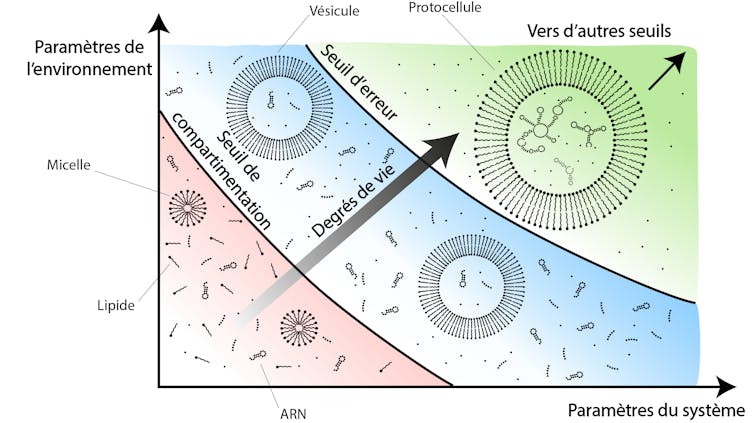
De nombreux seuils se cachent dans les transitions supposées de l’inerte au vivant. C’est, par exemple, le cas lorsqu’il s’agit d’imaginer l’apparition de compartiments ou de membranes. Ainsi, une solution micellaire de lipides exposée à une concentration en CO2 atmosphérique croissante donne lieu à la formation spontanée de vésicules lipidiques lorsqu’une valeur seuil de pression en CO2 est atteinte. Ces vésicules sont des compartiments microscopiques, constitués par une membrane de lipides, idéales pour la formation de protocellules. Cette transition de seuil joue ainsi un rôle clé pour comprendre l’apparition spontanée de ces structures essentielles pour penser l’avènement de cellules primitives.
Le « monde ARN »
Dans la recherche sur l’origine de la vie, un autre seuil attire tout particulièrement l’attention : le seuil d’erreur. Ce seuil délimite deux états dynamiques distincts dans lesquels peuvent se retrouver des systèmes de polymères réplicatifs : d’un côté la réplication est viable, de l’autre elle ne l’est pas, c’est-à-dire que l’information contenue dans les polymères ne se maintient pas dans le temps.
Les cellules des organismes vivants actuels contiennent de tels systèmes : les protéines permettent de répliquer l’ADN presque sans perte d’information. Dans le cadre de l’origine de la vie, certains supposent l’existence de systèmes d’ARN pouvant se répliquer les uns les autres. Cette molécule est très proche chimiquement de l’ADN, mais se présente sous la forme d’une simple hélice, contrairement à l’iconique double hélice de l’ADN.
Dans le cadre de l’hypothèse du « monde ARN », cette molécule est pressentie pour avoir pu apparaître avant l’ADN ainsi qu’avant les protéines, car elle peut stocker de l’information comme le premier et catalyser des réactions chimiques comme les secondes. L’ARN pourrait donc à lui seul soutenir les fonctions essentielles d’un système protovivant.
De même que tout système de polymères réplicatifs, la dynamique réplicative de systèmes d’ARN est régie par une transition de seuil d’erreur. Théoriquement, ce seuil d’erreur peut être franchi de plusieurs façons, notamment en jouant sur la taille des ARN répliqués et le taux d’erreur des ARN réplicatifs (aussi appelés réplicases) qui sont deux paramètres des systèmes en question. Ainsi, en faisant varier expérimentalement ces paramètres, il devrait être possible de faire émerger des systèmes d’ARN réplicatifs stables à partir d’une solution d’ARN, ce qui serait un pas de géant dans la compréhension de l’origine de la vie. Mais d’autres paramètres peuvent aussi être envisagés pour induire une telle transition de seuil, notamment des paramètres liés aux conditions de l’environnement comme la présence ou non de minéraux susceptibles de favoriser la production d’ARN, comme des argiles. Ainsi, une transition de seuil peut être de nature « intrinsèque » lorsqu’elle est régie par des paramètres de contrôle relatifs aux systèmes eux-mêmes, ou de nature « extrinsèque » quand elle repose sur des changements dans des paramètres environnementaux externes aux systèmes étudiés.
Dans d’autres cas, la transition d’un état dynamique à un autre peut relever de la fusion ou de l’intégration de plusieurs systèmes différents. Ainsi, une autre façon de franchir le seuil d’erreur consiste à introduire des compartiments dans lesquels viennent se réfugier des systèmes d’ARN réplicatifs, donnant alors naissance à des protocellules. C’est ce qu’on peut appeler des transitions « symbiotiques ».
L’articulation des seuils
La formalisation d’un seuil permet d’évaluer la plausibilité de ces différents modes de transition en fonction des caractéristiques des systèmes envisagés et de leur environnement supposé. Comme proposé par notre récente étude, une analyse approfondie du seuil d’erreur favorise la mise à jour d’une multitude de paramètres physiques, chimiques et géologiques, jouant sur la plausibilité du scénario de l’émergence d’ARN réplicatifs. Se focaliser sur un seuil en particulier permet ainsi d’aborder des aspects aussi éloignés que les compositions atmosphériques ou minéralogiques, les cycles de températures ou les dynamiques de compartimentation. Le concept de seuil se révèle être un outil interdisciplinaire privilégié pour faire des ponts entre des propositions théoriques sur l’origine de la vie et leurs réalisations pratiques.
Par ailleurs, articuler les seuils les uns par rapport aux autres permet d’aller encore plus loin en mettant à jour de possibles scénarios d’origine de la vie de plus en plus aboutis. Par exemple, nous avons vu qu’il existait un seuil au-delà duquel des vésicules lipidiques se formaient spontanément. Nous avons aussi vu que la présence de compartiments facilitait la transition vers des systèmes réplicatifs viables. Il est donc possible d’envisager un scénario où une solution contenant des ARN et des lipides franchirait ces deux seuils. L’élaboration de ce bref scénario, en deux transitions de seuils, permet une estimation de sa plausibilité sur la base de celles de ses seuils et incite à concevoir des environnements, voire des changements environnementaux, qui lui seraient plus ou moins favorables. De telles articulations sont particulièrement utiles pour dresser des chemins évolutifs complexes. L’intégration de la plausibilité de chacune des transitions prises en compte facilite l’estimation de la plausibilité de scénarios entiers et ainsi la comparaison entre eux.
On peut comprendre chaque transition de seuil, aussi minime soit-elle, comme une manière d’augmenter petit à petit le « degré de vie » d’un système physico-chimique. À la lumière de cette étude sur les seuils, la quantification du degré de vie de tout système situé quelque part entre l’inerte et le vivant pourrait se mesurer comme le ratio du nombre de seuils franchis sur le nombre de seuils envisagés dans un scénario d’origine de la vie donné. Même si nous ne connaissons pas encore ce chemin tortueux reliant l’inerte au vivant, la formalisation de seuils et de leurs transitions permet de concevoir une manière de s’approcher graduellement du vivant.
La Région Île-de-France finance des projets de recherche relevant de Domaines d’intérêt majeur et s’engage à travers le dispositif Paris Région Phd pour le développement du doctorat et de la formation par la recherche en cofinançant 100 contrats doctoraux d’ici 2022. Pour en savoir plus, visitez iledefrance.fr/education-recherche.