



Les engrais azotés forment un des piliers de la révolution agricole du XXe siècle, et sont devenus indispensables pour maintenir les forts rendements agricoles actuels.
Ces engrais sont fabriqués en puisant de l’azote présent sous forme de gaz dans l’air. L’azote gazeux étant une molécule chimique très stable, sa transformation en engrais nécessite énormément d’énergie, qui est apportée par l’utilisation d’hydrocarbures et participe donc à la production de gaz à effet de serre. De plus, les engrais apportés dans le sol sont souvent lessivés par les fortes pluies et entraînés dans les cours d’eau où ils facilitent la croissance d’algues étouffant les autres organismes vivants (phénomène d’eutrophisation).

Ainsi, le coût environnemental des engrais azotés – qu’ils soient industriels ou sous forme de lisier – est donc important, faisant de la réduction de leur utilisation une priorité à l’horizon 2030 pour l’agriculture mondiale.
Pourtant, certaines plantes peuvent se passer d’engrais azotés. Ces plantes appartiennent à quatre grands groupes botaniques, dont celui des légumineuses auquel appartiennent des plantes telles que les pois, haricots, lentilles, soja, arachide, mais aussi certains arbres (Acacia, Mimosa ou autres Robiniers). Comme lors du procédé chimique industriel, ces plantes puisent une grande partie de l’azote dont elles ont besoin directement dans l’air… grâce à la symbiose avec des bactéries qui vivent au niveau des racines.
Depuis de nombreuses années, les scientifiques se demandent comment cette association entre plantes et bactéries a pu évoluer. Si atteindre une telle compréhension pourrait peut-être, dans le futur, permettre de transférer à d’autres espèces la capacité à assimiler spontanément de l’azote – et ainsi limiter massivement l’utilisation d’engrais tout en maintenant des rendements élevés – la faisabilité d’une telle approche n’est pas aujourd’hui démontrée.
Dans notre récente étude publiée dans Nature Plants, nous avons analysé la manière avec laquelle plusieurs plantes légumineuses interagissent avec leurs bactéries symbiotiques, afin de retracer l’histoire évolutive de ces interactions.
Une symbiose au sein d’un organe dédié
Chez les plantes légumineuses, ce sont des bactéries vivant en symbiose avec ces plantes qui transforment l’azote gazeux en ammonium utilisable par la plante. Ce type de symbiose, dite « mutualiste », se retrouve de manière générale dans tout le vivant, et résulte de l’interaction entre deux organismes qui améliore réciproquement leur croissance et leur développement.
Cette symbiose a lieu au sein d’un organe spécifique au niveau de la racine de la plante, appelé « nodule », où les bactéries sont hébergées et fournissent de l’ammonium. Les partenaires y échangent aussi des signaux chimiques complexes.

Retracer l’origine des plantes symbiotiques nodulantes
Devant la diversité des plantes symbiotiques actuelles, il est difficile de savoir quelles caractéristiques physiologiques et génétiques sont nécessaires et suffisantes à la symbiose fixatrice d’azote. Distinguer les traits ancestraux de ceux qui sont apparus plus récemment chez les plantes symbiotiques devrait permettre d’établir la « recette génétique » de cette association.
En comparant les génomes de multiples plantes capables ou pas de réaliser la symbiose fixatrice d’azote, des travaux antérieurs ont montré que toutes les plantes symbiotiques possèdent un groupe de gènes communs, indispensables à cette association… Mais ces gènes sont aussi présents chez certaines plantes non-symbiotiques.
Lors de l’évolution, l’apparition de la symbiose ne serait donc pas liée à l’acquisition de nouveaux gènes mais plutôt à une modification de l’expression (ou activité) de ce groupe de gènes symbiotiques communs.

En effet, même si toutes les cellules d’un organisme possèdent les mêmes gènes, l’activité de ces derniers varie fortement d’une cellule à l’autre en réponse aux conditions environnementales (incluant, par exemple, la proximité de micro-organismes) et selon les stades de développement. Ainsi, les plantes légumineuses auraient acquis la capacité à réaliser la symbiose fixatrice d’azote en ré-utilisant des gènes impliqués dans différentes processus physiologiques (formation des racines latérales, interaction avec des champignons bénéfiques…) et en activant leur expression lors de l’interaction avec les bactéries symbiotiques.
C’est ce processus de « bricolage moléculaire », selon l’expression introduite par le biologiste François Jacob, que nous avons essayé de retracer.
Quand les plantes évoluent depuis un ancêtre commun grâce au « bricolage moléculaire »
Nous avons donc décidé de comparer les gènes qui sont spécifiquement activés dans les nodules de neuf espèces de plantes capables de former cette symbiose.
Plus précisément, notre objectif était d’identifier les gènes communément exprimés chez toutes ces espèces lors de la symbiose.
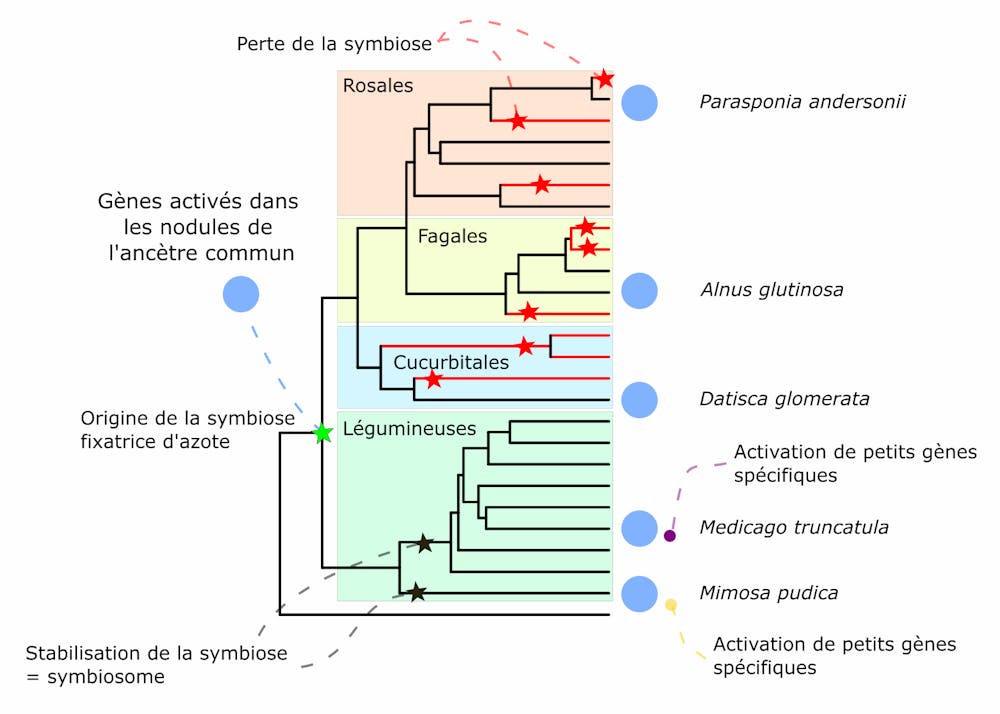
Nous avons observé que près d’un millier de gènes étaient exprimés de manière partagée dans les nodules des neuf plantes symbiotiques étudiées. L’explication la plus probable d’une telle similarité est que ces différentes espèces ont hérité de leur ancêtre commun, un « ancêtre symbiotique » qui vivait sur terre il y a environ 90 millions d’années, ce programme génétique permettant la formation et le fonctionnement des nodules.
Grâce aux connaissances acquises par ailleurs sur ces symbioses, nous avons pu identifier, dans cette liste, de nombreux gènes végétaux permettant aux plantes de percevoir les signaux chimiques produits par leurs bactéries symbiotiques, d’accueillir ces dernières dans leurs tissus, et de réaliser les processus moléculaires permettant de puiser l’azote de l’air.
Ainsi, « l’ancêtre symbiotique » était certainement capable de réaliser ces trois étapes indispensables au fonctionnement de la symbiose, via des mécanismes moléculaires reposant sur l’activité de ce groupe de gènes communs.
Des améliorations apparues indépendamment au fil de l’évolution
Mais l’évolution ne s’arrête jamais : certaines plantes descendant de cet ancêtre commun symbiotique ont perdu la capacité à réaliser cette symbiose. D’autres, au contraire, ont évolué des compétences symbiotiques particulières, des « ajustements » symbiotiques, leur permettant par exemple de réaliser la symbiose avec différents types de bactéries ou dans certaines conditions environnementales.
Nous nous sommes donc demandé si l’on pouvait déceler quand, dans l’évolution, avaient eut lieu ces ajustements.
Pour cela, nous nous sommes focalisés sur deux plantes (Mimosa pudica et Medicago truncatula) et avons étudié les gènes qui participent à la symbiose chez ces plantes sans toutefois être exprimés chez l’ancêtre commun. En effet, les plantes appartenant aux deux plus grandes familles de légumineuses (représentées par M. pudica et M. truncatula) ont très peu perdu leurs capacités symbiotiques et sont toujours capables de former des nodules aujourd’hui.
Il a récemment été proposé que la stabilité de cette capacité symbiotique au fil de l’évolution de ces plantes soit liée à la capacité de ces plantes à accueillir les bactéries à l’intérieur même des cellules végétales, au sein de structures appelées « symbiosomes ».
Nous disposons pour ces deux plantes d’une description précise de l’expression des gènes associée à chaque étape du processus de production des nodules, de formation des symbiosomes, et de fixation d’azote.
Nous nous sommes aperçu qu’un grand nombre de gènes associés à la formation des symbiosomes étaient spécifiques à chacune de ces deux espèces de plantes. En d’autres termes, ces gènes n’étaient pas présents dans le nodule de leur ancêtre commun, et la capacité à héberger les bactéries dans les cellules des nodules a donc émergé (évolué) indépendamment chez M. pudica et M. truncatula.
Ces « ajustements » symbiotiques auraient donc possiblement convergé vers un même mécanisme, le contrôle du symbiote, mais en utilisant différents processus moléculaires. Des travaux futurs devraient permettre de tester cette hypothèse.