

Contrairement à ce que l’on croit souvent, la recherche en écologie ne consiste pas seulement à observer la nature. Et certains chercheurs étudient des espèces et des écosystèmes virtuels qu’ils ont eux-mêmes créés. C’est ce qu’on appelle « faire de la modélisation ».
La modélisation en écologie consiste à reproduire des processus observés à l’aide d’équations mathématiques afin de comprendre le fonctionnement des systèmes étudiés. Ces équations représentent soit des flux, soit des variations d’abondance ou de biomasse d’espèces. Par exemple, la croissance d’une population de bactéries au cours du temps peut être représentée par une fonction exponentielle.
Les écologues peuvent également faire des prédictions sur l’état futur des écosystèmes.
Le processus de modélisation suit alors trois phases principales : l’observation, la compréhension et la prédiction. Cependant, il est parfois possible de comprendre sans pouvoir prédire, ou de prédire sans comprendre. Les avancées technologiques ont permis de développer des modèles complexes nécessitant des ordinateurs de plus en plus puissants qui permettent de lancer des simulations (prenant parfois plusieurs jours !).
Prévoir les effets des changements climatiques
La modélisation sert, entre autres, à comprendre les causes des changements climatiques et à prédire leur évolution. La distribution des espèces étant hautement dépendante du climat, si nous connaissons le climat futur et la distribution actuelle des espèces, nous devrions être capables de prédire une possible distribution future de celles-ci. C’est ainsi que nous pouvons imaginer la montée des espèces vers le nord.
Ces résultats sont très importants afin de préparer les gestionnaires aux changements climatiques. Ainsi, il est parfois suggéré aux forestiers de planter des essences d’arbres adaptées au climat futur. Il pourrait aussi être question de relocaliser certains vignobles, à l’image de la production de champagne désormais possible au Royaume-Uni.
Cependant, la présence d’une espèce dans un milieu ne dépend pas uniquement de l’environnement, mais aussi de ses interactions avec d’autres espèces. Par exemple, deux espèces végétales peuvent être en compétition pour l’accès à la lumière ou l’acquisition de nutriments. Dans ce cas, l’espèce la moins compétitrice risque de se faire supplanter. Ces espèces peuvent aussi être limitées dans leur expansion par une grande pression d’herbivorie ou une absence de pollinisateurs. Ajoutées aux modèles, ces variables les rendent plus complexes.
Observer la dynamique des espèces
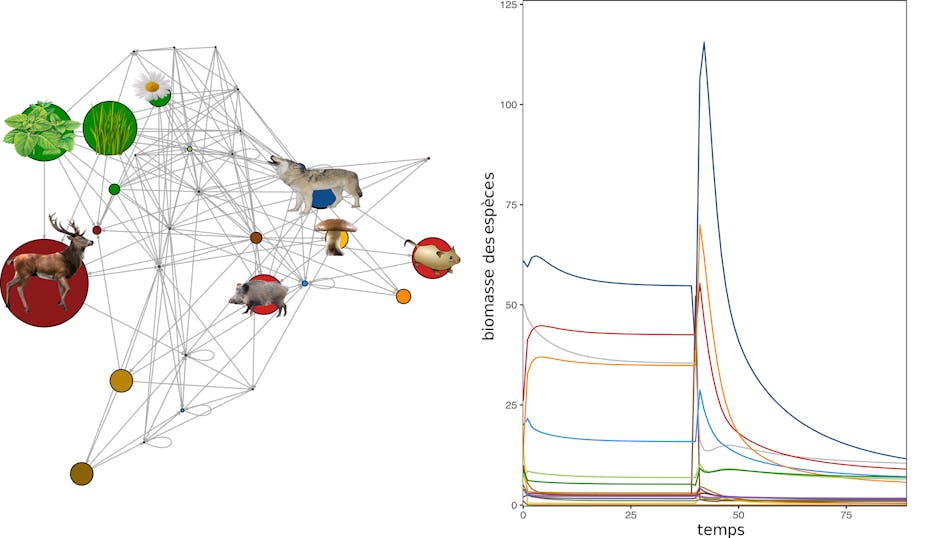
Dans la même optique, il est possible de modéliser les effets des perturbations sur des communautés entières comportant des végétaux, des herbivores et des carnivores. Une perturbation peut être progressive (comme un changement climatique) ou brutale (comme un feu ou une coupe forestière) et donc d’origine naturelle ou humaine (anthropique). Les perturbations favorisent certaines espèces (parfois des espèces exotiques et/ou pathogènes) et peuvent en éliminer d’autres.
En connaissant les interactions entre espèces, des prédictions peuvent être émises quant à l’effet d’une perturbation sur une communauté entière. Dans les modèles de réseaux trophiques (un ensemble de chaînes alimentaires) où seulement les interactions consommateurs-ressources sont prises en compte, chaque espèce consomme des ressources à des taux variables, qui sont souvent déterminés à partir de sa masse corporelle. Des espèces peuvent donc être retirées ou ajoutées aux modèles et les effets de ces modifications sur les autres espèces peuvent être calculées.
La modélisation de la dynamique de communautés d’espèces a permis de révéler des effets en cascade difficilement observables sur le terrain et difficilement prédictibles. Par exemple, la modélisation d’une grande pression de pêche sur les poissons carnivores et/ou de grande taille a montré que la pêche modifie les communautés marines dans leur ensemble, au niveau de leur composition en espèces et de leur fonctionnement. C’est à cause de cette pression de pêche que l’on a pu, par exemple, observer ces dernières années une surabondance de méduses en Méditerranée, conséquence de la surpêche de leurs prédateurs. Cela a entraîné, en cascade, une diminution des proies des méduses telles que les petits poissons.
Simplifier la réalité
Les modèles peuvent être très réalistes ou très abstraits en fonction de leurs objectifs. Certaines théories écologiques ont ainsi été développées en utilisant des espèces imaginaires. Malgré cela, ces théories ont été confirmées a posteriori par des observations. La théorie de la biogéographie des îles, par exemple, permet de comprendre comment les flux d’espèces entre îles et continents façonnent les populations insulaires.

{kind=link}
De nos jours, cette théorie élaborée mathématiquement est communément utilisée dans la gestion des écosystèmes. D’autres modèles sont construits et paramétrés suivant de vraies communautés. Par exemple, les communautés de micro-organismes présents dans les feuilles de forme tubulaire des Sarracénies pourpres sont utilisées pour comprendre, entre autres, comment les interactions entre espèces affectent le fonctionnement des communautés.
Il existe un gradient dans le réalisme des modèles. S’ils s’avèrent trop abstraits, cela peut induire du scepticisme quant à la justesse de leurs résultats. C’est pourquoi il est important de se rappeler qu’un modèle est construit dans le but de simplifier la réalité afin de comprendre des mécanismes précis. Un modèle comporte donc uniquement les variables d’intérêt pour une question particulière, et, comme l’a déclaré le statisticien George Box, « tous les modèles sont faux, mais certains sont utiles ».
C’est justement pour répondre à des questions diverses que de nombreux modèles, à première vue similaires, sont construits de manière un peu différente. C’est alors l’ensemble de ces modèles qui permet de comprendre différents aspects du fonctionnement des communautés et des écosystèmes, et de prédire leur réponse aux changements environnementaux.
Modélisation et recherche empirique
Bien que les modèles en écologie soient maintenant des outils indispensables pour comprendre le monde qui nous entoure, leur simplicité ne permet pas de saisir entièrement la complexité que l’on trouve dans la nature. Cette complexité, résultant d’un grand nombre d’espèces, d’interactions et de facteurs environnementaux, rend cependant la compréhension globale des systèmes biologiques impossible si l’on ne prend en compte que les expériences empiriques.
La modélisation reste donc indissociable des expériences de terrain. La récolte de données alimente ainsi le fonctionnement des modèles, lesquels offrent des prédictions dont la justesse peut être vérifiée empiriquement. L’utilisation conjointe en écologie de la modélisation et de la recherche empirique ouvre ainsi les portes à une nouvelle ère, à de nouvelles réponses et, bien sûr, à de nouvelles questions.
Stéphanie Préfontaine et Madelaine Proulx ont participé à la rédaction de cet article.