

When the young naturalist Alfred R. Wallace left England to explore the New World in 1848, one of his key observations in the Amazon region was that large rivers were unsurpassable barriers for some species of animals, particularly primates and birds. Even more interesting, he noticed that closely related species often occupied forests on opposite sides of a river, but they were never found together.
Indeed, there are hundreds of examples of cross-river pairs among Amazonian birds, a phenomenon not seen anywhere else on Earth. Nearly one-third of the approximately 240 bird species that normally inhabit the forests on one side of the lower Rio Negro in Brazil are replaced by a closely related species on the other side. For example, on the left bank you will see Black-spotted Barbets (Capito niger) with red throats. On the right bank you will only see the closely related Guilded Barbets (Capito auratus) with orange throats.

Amazonian rivers have fascinated me since I first visited the region in 1999 and have been the main focus of my research as a tropical biologist. I was captivated by the replacement patterns Wallace described, and spent countless hours studying avian distribution maps.
Despite years of study, however, evolutionary biologists remain uncertain of the role rivers have played in the speciation process in Amazonia. Do rivers generate new species? Or simply act as secondary barriers, providing natural boundaries for species formed elsewhere? Did all these paired lineages diverge at the same time, divided by a common barrier? Or did each species follow its own evolutionary path?
These were the questions that guided my colleagues and me when we set out to explore the Rio Negro, still one of the most pristine regions of Amazonia. Like expeditions conducted by earlier naturalists, ours involved long boat journeys to reach remote locations. But we were armed with computers, digital imagery, GPS devices, digital audio recorders and liquid nitrogen to keep our samples at very low temperatures until we could perform genetic analysis back in the lab. Our plan was to use tissue samples from birds on opposite sides of the river to assess their evolutionary history.
Evolving from one species to two
By looking at the amount of genetic differentiation between individuals on different banks, modern naturalists are able to track down approximately how much time has passed since these populations began their independent evolutionary histories. What biologists cannot agree upon is what role these rivers played in the evolutionary history of the species they currently divide.
One obvious possibility is that the range of the ancestral species was dissected by a newly formed river, isolating big chunks of forests. Cutoff populations would slowly change and differentiate from one another. Given enough time, their differences would become great enough that they’d no longer recognize each other as potential mates with which to pair and raise offspring – they’d become different species. This idea became known as the riverine barrier hypothesis. It’s the oldest explanation for why there are so many species in Amazonia.
An alternative model suggests that rivers act as secondary barriers. According to this idea, although they’re currently important to define species’ geographical limits, rivers had nothing to do with the initial separation of that ancestral population.
It’s hard not to ask the most obvious question here, though. Birds engage in epic long-distance migrations and are able to travel from pole to pole. How in the world could an Amazonian river, even a large one, represent much of a barrier? Can’t birds simply fly across the river, reunite with their relatives and avoid becoming two different species?
The short answer is, apparently not. For many forest-dwelling species, rivers really do seem to be insurmountable barriers. Experiments suggest that many bird species are not capable of flying even a hundred yards over open landscape, let alone crossing several kilometers of a mighty Amazonian river.

Testing the role of rivers in avian evolution
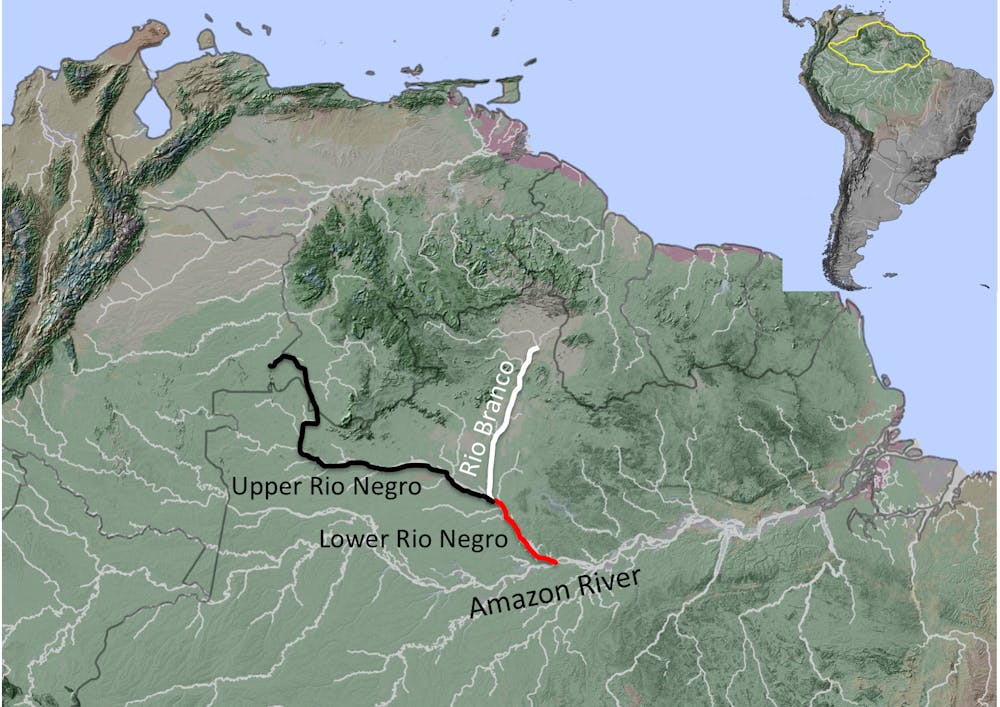
Our first step was to map the exact location of these pair replacements upriver. All the pairs studied were known to turnover across the lower Rio Negro, which ranges as wide as 10 kilometers in some places. Upriver, however, the river is much narrower and replacement patterns become more complicated, involving minor rivers.
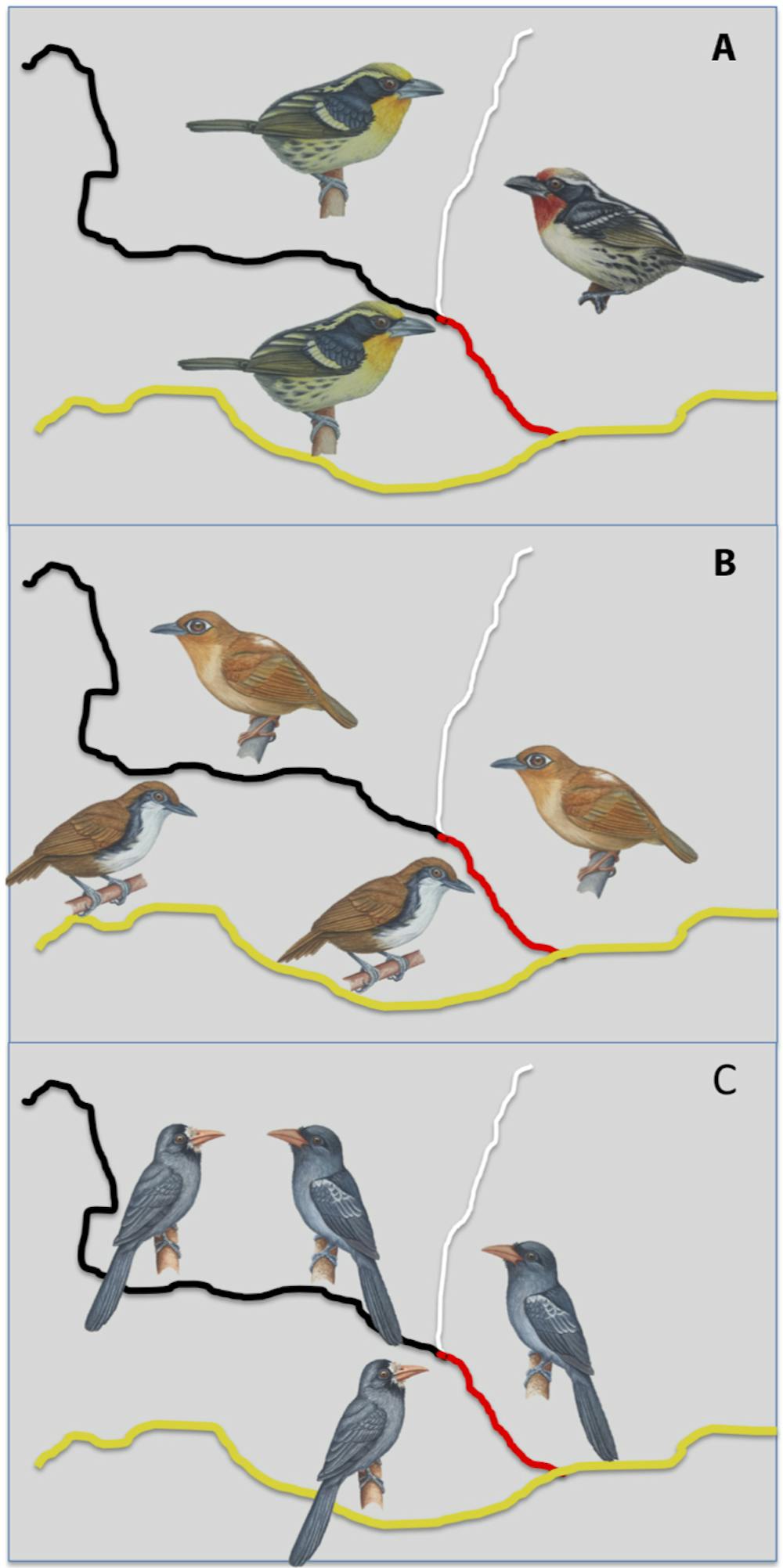
When differences between populations are relatively large – like plumage patterns, colors or song – pairs are often considered different species. When differences are subtle – such as size or feather hue or tone – ornithologists tend to consider them different subspecies. Whether pairs of species or pairs of subspecies, biologists refer to them as “pairs of taxa.”
We investigated in detail 74 pairs of taxa whose ranges were divided by various combinations of the Rio Negro and its largest tributary, the Rio Branco. These two rivers are both biogeographical barriers for dozens of avian species. After more than a decade of fieldwork, and with the contribution of major Brazilian and U.S. museums and collections, our team had obtained distributional and genetic data for almost all bird species and subspecies that differed on either side of the river.
We reasoned that if a river dissected the landscape and separated many avian populations at the same time, pairs should present roughly similar times of divergence. If rivers acted as secondary barriers, pairs will likely present a whole suite of ages. In this case, pairs could be older than the rivers that currently bound their distributions, since according to this model, the original division of a species in two doesn’t depend on the emergence of the river.
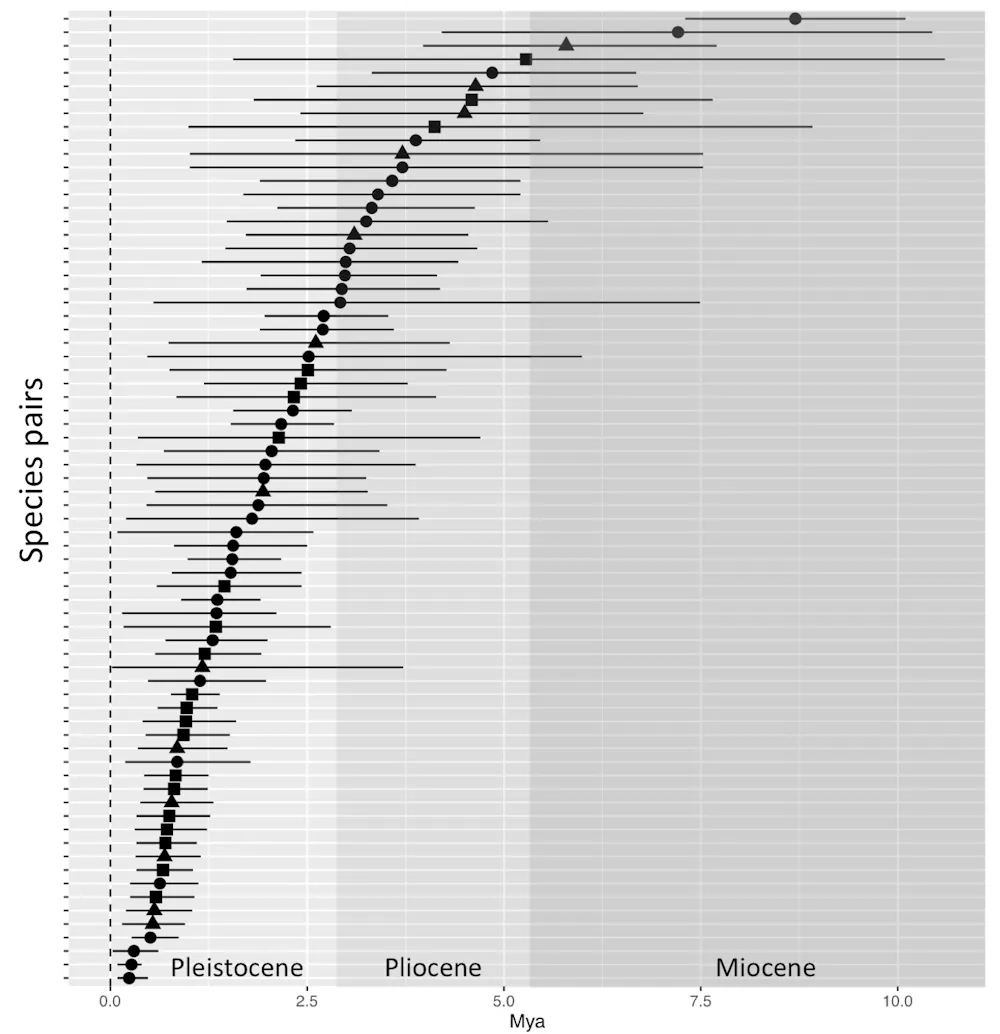
We used time-calibrated molecular data to figure out approximately when each of our 74 pairs of related birds went their separate ways, evolutionarily. Some genes mutate at predictable, steady rates, allowing scientists to estimate the time that’s elapsed since any given pair of organisms diverged. The more changes in their genomes, the longer it’s been since they shared a common ancestor. It’s like looking at what researchers call a “molecular clock.”

When we looked at the molecular clocks of our bird samples, we found that divergence events were not clustered within a particular time frame. Instead they ranged from 0.2 to 8 million years ago. So it’s unlikely that all avian pairs currently divided by a common barrier, such as the Negro or the Branco, were generated by the genesis of those rivers.
Furthermore, geomorphological data suggest that these rivers established their current positions relatively recently. Approximate dates for the Rio Negro are around a million years, whereas the Rio Branco is apparently much more recent, around 20,000 years. Therefore, both rivers seem to be much younger than most bird pairs they currently divide, supporting the secondary contact barrier model: The rivers today maintain a boundary, but the timing suggests they couldn’t have been responsible for initially separating most of the avian pairs we studied.
On the other hand, the Rio Negro does appear to have a million-year history here. Our models cannot reject a common diversification event happening at around that age for 12 pairs of avian taxa, whose DNA indicated they’d diverged within the last million years. So while the origin of most pairs studied is likely not related to the genesis of the rivers, it is possible that the Rio Negro, in particular, represented a primary barrier for some species.
Our new study, published in Science Advances, offers compelling evidence that Amazonian rivers may hold a dual evolutionary role. They can act as primary barriers for some lineages, as proposed by the riverine barrier hypothesis. But more frequently, they act as secondary barriers for most avian lineages. These results come from only one, albeit important, Amazonian region, and similar studies from other basins will place our results into a broader context.