
The animal world has many weird and wonderful ways of having sex. Some animals, such as snails, are hermaphrodites - able to make both eggs and sperm simultaneously. Some, such as wrasses and parrot-fish, initially hatch as male but transition to female in later life as they get older. Still others, including some lizard species, have dispensed with males entirely, and the females reproduce by parthenogenesis – laying fully fertile eggs without the need for sperm.
But what of the majority of animal species – the ones that are born either male or female, and then stay that way throughout their life history? Surely that has to be a simpler story? Apparently not. Even in these comparatively straightforward cases, there are still complex trade-offs that govern how, when and why animals develop as male or female.
Hitherto it has been believed that this choice is determined either by the genes passed down from the parents, as in mammals with X and Y chromosomes; or by environmental factors, as in turtles where sex determination depends on the incubation temperature of the egg. But new research shows that it is more complex still. Contrary to prediction, embryonic turtles can control their own development and choose for themselves whether to become a boy or a girl.

Why sex ratio matters
The goal of all life is not just to reproduce, but to ensure continued reproduction for generations to come: to maximise the future reproductive success of one’s offspring. A simple prediction is that this causes sex ratios to converge on a stable 1:1 mix of males and females. This is the so-called “Fisherian” sex ratio.
Simply put, if there are more females than males in the population, then genes which favour the production of more male offspring will be favoured by evolution – and vice versa – since it will be easier for offspring of the rarer sex to find mates. This means that natural selection should in principle ensure that offspring sex ratios remain equal or near-equal.
But biology is never quite that simple. Another important factor is the physical and social status of the resulting offspring. While males and females will on average have exactly the same number of offspring, female reproductive capacity is more predictable than male reproductive capacity.
For example, a single bull elephant seal can control a harem of up to 100 cows and sire 100 calves in a year, while the other 99 males produce no offspring at all. Meanwhile, the majority of the cows on the beach sire a single calf each.
From the standpoint of maximising the number of descendants, this means that having daughters is like investing in gilts – the return on investment is not much to shout about, but it’s guaranteed. Having sons, on the other hand, is like playing the lottery – most of the time there’s no return, but a small percentage will win the jackpot.

Mother knows best?
This is where the games begin. If the parents can predict the outcome of this lottery – if they can gauge their future offspring’s likely reproductive success - then they have an incentive to try and skew their offspring sex ratio away from 1:1.
For example, if the mother is in poor physical condition, it’s very likely that her offspring will be smaller, less fit, generally runtier and unlikely to compete successfully for mates. Under those circumstances, it makes sense for her to “play it safe” and bear more daughters, since that virtually guarantees at least a few grandchildren. But, if the mother is in the prime of health, her offspring are likely to in turn be stronger and therefore more attractive to potential mates, and so she can “shoot for the moon” by bearing more male offspring.
Sex skewing can be achieved in a number of different ways – by varying which chromosomes are passed on in species with chromosomal sex determination, or by varying incubation temperature in species with temperature-dependent sex determination, or even by selective reabsorption of embryos of the “unwanted” sex in species such as mammals that develop internally.
This prediction, first made by Bob Trivers and Dan Willard in 1973, has since been shown to hold true in at least some animal species, including insects, birds and mammals. In some cases, it is the father’s physical condition that makes the difference, with healthier fathers bearing more sons.
But robust evidence for adaptive sex ratio skewing remains to be established for most species due to a variety of other confounding factors such as seasonality in offspring sex ratios that make it very challenging to measure such effects reliably.
Go your own way
Scientists have long presumed that the embryo is merely a passive participant in this process: that any decision-making comes entirely from the parental side. But the new paper by a team from the Institute of Zoology in the Chinese Academy of Sciences shows, for the first time, that the embryo has some measure of control over its destiny. Each embryo, it seems, can have its own “interests” and make its own decision over which sex to be.
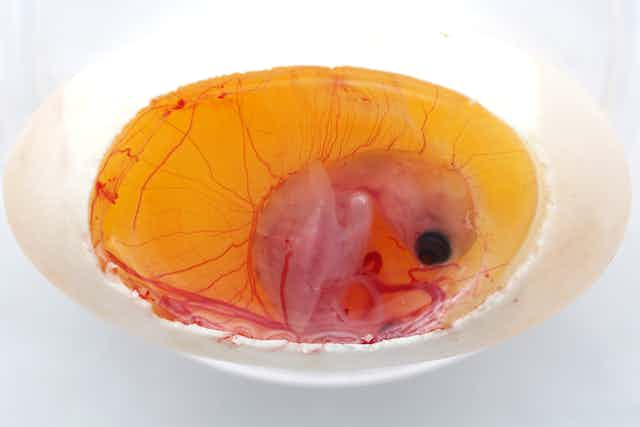
Working in freshwater turtles (Mauremys reevesii), a species in which sex is determined by the temperature at which the egg is incubated, they confirmed that incubation at cool temperatures triggers male development, while incubation at warmer temperatures triggers female development. Next, they showed that each individual egg does not have a single fixed temperature but can have hot spots and cool spots.

Finally, in a range of experiments designed to mimic different environmental conditions, they found that embryos preferentially chose to locate themselves at the warmer end of the egg when incubated in cool conditions, and at the cooler end when incubated in warm conditions. This tended to normalise sex ratios back towards the Fisherian 1:1 ratio. Injecting eggs with a drug that blocked the embryo’s temperature sensors interrupted this process and led to increasingly skewed sex ratios.
A lot still remains to be seen, in particular the extent to which the embryos are choosing their sex, rather than choosing a preferred temperature for other reasons unrelated to sex determination. It would be helpful, for example, to investigate whether the embryo behaviour accords with the Trivers-Willard theory that increasing embryo robustness should lead to increased numbers of male offspring. For example, whether smaller, slower-developing embryos (or embryos from mothers in poor condition) preferentially choose a female fate, and more robust embryos preferentially choose a male fate.
But what is certain is that how, when and why animals develop as male or female is much more complex than we ever imagined.